
Dr Mark Bass
School of Biosciences
Lecturer


- Profile
-
- 2025-present: University of Sheffield Lecturer, School of Biosciences, University of Sheffield, UK
- 2015-2025: University of Sheffield Lecturer & Deputy Director of Undergraduate Student Recruitment, School of Biosciences, University of Sheffield, UK
- 2009-2015: Wellcome Trust Research Career Development Fellow, School of Biochemistry, University of Bristol, UK
- 2001-2009: Postdoctoral Research Associate, Faculty of Life Sciences, University of Manchester, UK
- 1997-2001: PhD, Biochemistry, School of Biological Sciences, University of Leicester, UK
- 1996-1997: MRes, Biochemistry University of Leicester, UK
- 1993-1996: BSc (Hons), Biochemistry University of Leicester, UK
- Research interests
-
Fibroblast migration during wound healing: signalling from extracellular matrix receptors to Rho-family GTPases
Healing defects are one of the largest current health challenges, with chronic wounds frequently requiring amputation of the affected limb. The National Health Service expends £5.3 billion, 4% of the NHS budget, annually, treating 2.2 million patients for wounds, with £1.94 billion alone spent on treating chronic wounds such as venous leg ulcers. The challenge continues to escalate as risk factors such as age and diabetes rise, making wound management one of the major challenges of modern medicine.
Upon wounding healthy skin, inflammatory cells combat infection, fibroblasts migrate into the wound bed and contract the defect, and finally re-epithelialisation closes the gap. However, these processes become less efficient with age and risk factors such as diabetes, obesity or smoking, eventually leading to the formation of chronic wounds that include pressure ulcers, venous leg ulcers and diabetic foot ulcers. The two hallmarks of a chronic wound are a chronic inflammatory response as the skin tries unsuccessfully to deal with infection and failure by fibroblasts to proliferate and migrate in response to stimuli such as growth factors. The fact that scars are usually a fraction of the size of the original wound demonstrates very simply the importance of fibroblast migration and wound contraction, and improving this process is the core objective of our work.
Our laboratory investigates the activation of fibroblasts upon wounding, and the mechanisms by which migration is directed by regulation of the Rho family GTPases, especially the protrusion regulator, Rac1. Our work ranges from the investigation of signalling networks in single cells, through the analysis of in vivo wound healing models, to the development of therapies for treatment of patients.
The work can be divided into three main areas:
1) Regulation of membrane protrusion by Rho-family GTPases
Cell migration requires cycles of protrusion at the leading edge, adhesion to an extracellular matrix and contraction within the cell body. These events are driven by cycles of activation of Rac1 and RhoA. The appearance of fibronectin in wounded tissue triggers cycles of Rac1 and RhoA activity in fibroblasts by engagement of the fibronectin sensor, syndecan-4.
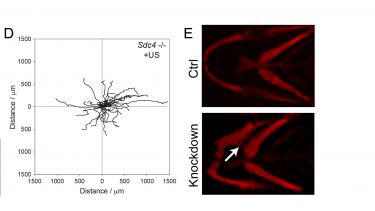
We are examining how syndecan-4 regulates and coordinates Rac1 and RhoA signals by combining traditional biochemistry with live cell imaging techniques. We have developed artificial skin dermis (A) that we can use as a substate to analyse signalling and behaviour by individual cells. For example, we can study GTPase activation using FRET techniques (B) and trafficking of GTPases using photoactivation and time-lapse microscopy (C). Perturbation of components of the syndecan-4 signalling chain by RNAi is revealing that syndecan-4 synchronises the activation/inhibition of Rac1 and RhoA signals and more importantly localises protrusion. The functional consequences of these signals can be tested by examining the migration of cells through the complex fibrillar matrices that are structurally similar to skin (D). We find that sydecan-4-directed GTPase regulation is necessary for persistent migration along matrix fibres. We can go even further by demonstrating that inhibition of the signals causes defects in zebrafish development (E), demonstrating broad effects on mesenchymal migration.
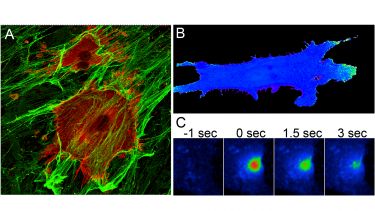
A) Fibroblasts stained for focal adhesion markers (red) embed into a 3D fibrous matrix (green).
B) Active Rac1 (green) is polarised to the front of a migrating cell, detected by FRET.
C) Photoactivation of fluorophore-labelled Rac1 allows analysis of the redistribution of Rac1 over time in response to different stimuli.
D) Migration tracks of fibroblasts migrating through a 3D fibrous matrix, demonstrating both speed and directionality of migration.
E) Knockdown of molecules that regulate trafficking of Rac1 leads to misalignment of pharyngeal arches in the developing zebrafish, due to compromised neural crest migration.
2) Cooperation between transmembrane receptors regulates focal adhesion dynamics
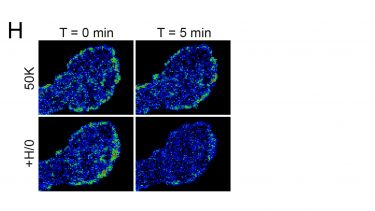
Membrane protrusion must be coordinated with formation and dissolution of focal adhesions for migration to occur. Furthermore, crosstalk between different transmembrane receptors leads to a coordinated response to growth factor and extracellular matrix signals. We are investigating the regulation of integrin trafficking by syndecan-4 using atomic force microscopy to measure adhesive strength (F+G) and TIRF to follow removal of β1-integrin from the adhesion plane upon engagement of syndecan-4 (H). By extrapolating the effect of syndecan-4 on adhesion receptors to investigate the effects on growth factor receptor activation and trafficking, we gain a fuller picture of how signals are coordinated. Finally, we are also using mass spectrometry to identify key trafficking regulators, an approach that is likely to identify new therapeutic targets for the promotion of fibroblast migration and healing.
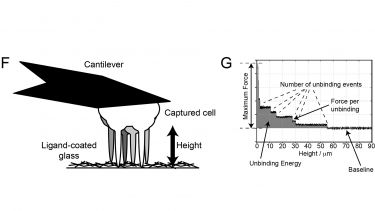
F) Single cells captured on the cantilever of an atomic force microscope can be brought into contact with an extracellular matrix before withdrawal of the cell to measure strength of adhesion.
G) Measurements of cantilever deflection in the atomic force microscope allow the force required to detach a cell to be calculated, and the effects of specific molecules on cell avidity to be determined.
H) Imaging of GFP-β1-integrin in the adhesion plane by TIRF reveals that syndecan-4 engagement triggers internalisation of integrin.
3) Ultrasonic healing therapies
The translation of our findings to in vivo healing models, and subsequently patient therapies is a crucial aspect of our work. Disruptions in signalling by transmembrane receptors or Rac1 cause migration to become less efficient and lead to delays in healing processes. For example, deletion of the transmembrane receptor, syndecan-4, is not lethal, but does delay skin healing (I+J). Introduction of pathological risk factors, such as age or diabetes cause similar delays in mice.
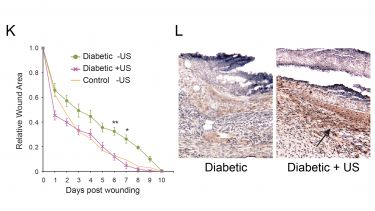
We are investigating techniques to reverse healing defects by activating Rac1. Our most notable advances have come from the use of low-intensity pulsed ultrasound to stimulate fibroblast migration. (K) We find that skin wounds heal more slowly in diabetic (green curve) than healthy mice (orange curve). However, normal healing can be restored in diabetic animals by daily treatments with ultrasound (pink line). The effect of ultrasound can be seen at the cellular level in biopsies as the number of brown fibroblasts recruited to an ultrasound-treated diabetic wound far exceeds that recruited to an untreated wound (L).
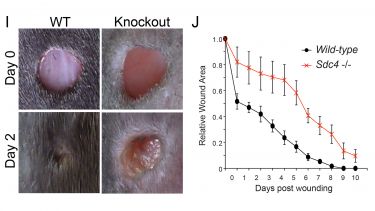
I) Knockout of syndecan-4 causes a defect in contraction of skin wounds. J) Knockout of syndecan-4 (red line) causes a delay in the closure rate of skin wounds.
K) The healing defects of diabetic mice (green) can be restored to the rates of healthy control mice (orange) by the application of ultrasound (pink).
L) Ultrasound treatment stimulates the recruitment of fibroblasts (brown) to the wounds of diabetic animals.
By combining this range of approaches to address the mechanism of fibroblast migration during wound healing, we move closer to developing some promising therapies, and bringing them into mainstream clinical use.






- Publications
-
Show: Featured publications All publications
Featured publications
Journal articles
- Inhibition of EphA2 by syndecan-4 in wounded skin regulates clustering of fibroblasts. Journal of Molecular Cell Biology, 17(1). View this article in WRRO


- The β3‐integrin endothelial adhesome regulates microtubule‐dependent cell migration. EMBO reports, 19(7).
- Ultrasonic Stimulation of Mouse Skin Reverses the Healing Delays in Diabetes and Aging by Activation of Rac1. Journal of Investigative Dermatology, 135(11), 2842-2851. View this article in WRRO
- Coronin-1C Protein and Caveolin Protein Provide Constitutive and Inducible Mechanisms of Rac1 Protein Trafficking. JOURNAL OF BIOLOGICAL CHEMISTRY, 290(25), 15437-15449. View this article in WRRO
- Coronin-1C and RCC2 guide mesenchymal migration by trafficking Rac1 and controlling GEF exposure. Journal of Cell Science , 127(19), 4292-4307. View this article in WRRO
- Resolution Mediator Chemerin15 Reprograms the Wound Microenvironment to Promote Repair and Reduce Scarring. Current Biology, 24(12), 1406-1414. View this article in WRRO
- Syndecan-4 phosphorylation is a control point for integrin recycling. Developmental Cell, 24(5), 472-485. View this article in WRRO
- A Syndecan-4 Hair Trigger Initiates Wound Healing through Caveolin- and RhoG-Regulated Integrin Endocytosis. Developmental Cell, 23(5), 1081-1082.
- Proteomic Analysis of Integrin Adhesion ComplexesA presentation from the 6th British Society for Proteome Research (BSPR)–European Bioinformatics Institute (EBI) Meeting “Multiscale Proteomics: From Cells to Organisms” at the Wellcome Trust Conference Centre, Cambridge, UK, 14 to 16 July 2009. The Presentation also complements the
Science Signaling
Research Article by Humphries
et al.
published 8 September 2009.. Science Signaling, 4(167).
- Therapeutic Ultrasound Bypasses Canonical Syndecan-4 Signaling to Activate Rac1. Journal of Biological Chemistry, 284(13), 8898-8909.
- p190RhoGAP is the convergence point of adhesion signals from alpha 5 beta 1 integrin and syndecan-4.. Journal of Cell Biology, 181(6), 1013-1026. View this article in WRRO
- Syndecan-4-dependent Rac1 regulation determines directional migration in response to the extracellular matrix.. Journal of Cell Biology, 177(3), 527-538. View this article in WRRO
All publications
Journal articles
- Inhibition of EphA2 by syndecan-4 in wounded skin regulates clustering of fibroblasts. Journal of Molecular Cell Biology, 17(1). View this article in WRRO
- The β3‐integrin endothelial adhesome regulates microtubule‐dependent cell migration. EMBO reports, 19(7).
- Chlorhexidine hexametaphosphate as a wound care material coating: antimicrobial efficacy, toxicity and effect on healing. Nanomedicine, 11(16), 2049-2057.
- Ultrasonic Stimulation of Mouse Skin Reverses the Healing Delays in Diabetes and Aging by Activation of Rac1. Journal of Investigative Dermatology, 135(11), 2842-2851. View this article in WRRO
- Comparing the Affinity of GTPase-binding Proteins using Competition Assays. Journal of Visualized Experiments(104).
- Coronin-1C Protein and Caveolin Protein Provide Constitutive and Inducible Mechanisms of Rac1 Protein Trafficking. JOURNAL OF BIOLOGICAL CHEMISTRY, 290(25), 15437-15449. View this article in WRRO
- Coronin-1C and RCC2 guide mesenchymal migration by trafficking Rac1 and controlling GEF exposure. Journal of Cell Science , 127(19), 4292-4307. View this article in WRRO
- Resolution Mediator Chemerin15 Reprograms the Wound Microenvironment to Promote Repair and Reduce Scarring. Current Biology, 24(12), 1406-1414. View this article in WRRO
- Resolution Mediator Chemerin15 Reprograms the Wound Microenvironment to Promote Repair and Reduce Scarring. Current Biology, 24(12), 1435-1435.
- RhoG Protein Regulates Platelet Granule Secretion and Thrombus Formation in Mice. Journal of Biological Chemistry, 288(47), 34217-34229.
- Syndecan-4 phosphorylation is a control point for integrin recycling. Developmental Cell, 24(5), 472-485. View this article in WRRO
- A Syndecan-4 Hair Trigger Initiates Wound Healing through Caveolin- and RhoG-Regulated Integrin Endocytosis. Developmental Cell, 23(5), 1081-1082.
- Syndecan and integrin interactomes: large complexes in small spaces. Current Opinion in Structural Biology, 22(5), 583-590.
- Induction of adhesion-dependent signals using low-intensity ultrasound.. Journal of Visualized Experiments(63). View this article in WRRO
- SNX17 protects integrins from degradation by sorting between lysosomal and recycling pathways.. Journal of Cell Biology, 197(2), 219-230. View this article in WRRO
- Syndecan-4 independently regulates multiple small GTPases to promote fibroblast migration during wound healing.. Small GTPases, 3(2), 73-79. View this article in WRRO
- SHARPINing integrin inhibition. Nature Cell Biology, 13(11), 1292-1293.
- A Syndecan-4 Hair Trigger Initiates Wound Healing through Caveolin- and RhoG-Regulated Integrin Endocytosis. Developmental Cell, 21(4), 681-693.
- Proteomic Analysis of Integrin Adhesion ComplexesA presentation from the 6th British Society for Proteome Research (BSPR)–European Bioinformatics Institute (EBI) Meeting “Multiscale Proteomics: From Cells to Organisms” at the Wellcome Trust Conference Centre, Cambridge, UK, 14 to 16 July 2009. The Presentation also complements the
Science Signaling
Research Article by Humphries
et al.
published 8 September 2009.. Science Signaling, 4(167).
- Proteomic Analysis of Integrin-Associated Complexes Identifies RCC2 as a Dual Regulator of Rac1 and Arf6. Science Signaling, 2(87).
- Giving off mixed signals—Distinct functions of α5β1 and αvβ3 integrins in regulating cell behaviour. IUBMB Life, 61(7), 731-738.
- An integrin-α4–14-3-3ζ–paxillin ternary complex mediates localised Cdc42 activity and accelerates cell migration. Journal of Cell Science, 122(10), 1654-1664.
- Syndecans Shed Their Reputation as Inert Molecules. Science Signaling, 2(64).
- Therapeutic Ultrasound Bypasses Canonical Syndecan-4 Signaling to Activate Rac1. Journal of Biological Chemistry, 284(13), 8898-8909.
- p190RhoGAP is the convergence point of adhesion signals from alpha 5 beta 1 integrin and syndecan-4.. Journal of Cell Biology, 181(6), 1013-1026. View this article in WRRO
- Synergistic control of cell adhesion by integrins and syndecans. Nature Reviews Molecular Cell Biology, 8(12), 957-969.
- Syndecan-4-dependent Rac1 regulation determines directional migration in response to the extracellular matrix.. Journal of Cell Biology, 177(3), 527-538. View this article in WRRO
- Integrins and syndecan-4 make distinct, but critical, contributions to adhesion contact formation. Soft Matter, 3(3), 372-372.
- Cytoplasmic interactions of syndecan-4 orchestrate adhesion receptor and growth factor receptor signalling. Biochemical Journal, 368(1), 1-15.
- Further characterization of the interaction between the cytoskeletal proteins talin and vinculin. Biochemical Journal, 362(3), 761-761.
- Talin contains three similar vinculin-binding sites predicted to form an amphipathic helix. Biochemical Journal, 341(2), 257-263.
- Integration of the Rac1- and actin-binding properties of Coronin-1C. Small GTPases, 6(1). View this article in WRRO
Conference proceedings
- Integrin‐Syndecan Cooperation Governs the Assembly of Signalling Complexes during Cell Spreading (pp 178-192)
Preprints
- The β3-integrin endothelial adhesome regulates microtubule dependent cell migration, Cold Spring Harbor Laboratory.
- Inhibition of EphA2 by syndecan-4 in wounded skin regulates clustering of fibroblasts. Journal of Molecular Cell Biology, 17(1). View this article in WRRO
- Teaching activities
-
Undergraduate:
- BMS106 Pathobiology
- BMS109 Cell & Molecular
- BMS242/243 Research Career Paths
- BMS336 Modelling Human Disease and Dysfunction
- Level 3 Final Year Dissertation Modules (Coordinator)
- Professional activities and memberships
-
Summer Research Experience Scheme (Coordinator)